山口大学 農学部 生物機能科学科
応用微生物学研究室
Laboratory of Applied Microbiology, Yamaguchi University
〒753-8515 山口市吉田1677-1 山口大学農学部生物機能科学科 応用微生物学研究室
Tel: (083) 933-5857; Fax: 5820; Email: obi(at)yamaguchi-u.ac.jp
English
Research Subject
酸化発酵の生理学的研究とその産業利用
酢酸菌は、高濃度の糖やアルコールを含む花蜜・果実やその酸敗した果実酒などの中で様々な
微生物と共存・競合して生育しており、この生育環境の中で極めて特異な生育生理を示します。
つまり、酢酸菌は特定の糖やアルコールを利用しにくい糖酸(有機酸)に急速に酸化変換し、
培地中に高濃度に蓄積することができます。この「酸化発酵」と呼ばれる反応は、共存する微
生物種による糖・アルコールの利用を妨害すると同時に、それら微生物の生存に不向きな
(有害な)有機酸の蓄積と低pH環境をもたらします。しかし、これらの条件では、酢酸菌自身
の生育も決してよいものではありません。しかしながら、酢酸菌はそれらの有機酸や低pHに比
較的耐性で、しかも一定時間後にそれらの糖酸を資化・利用して再び生育を開始することがで
きる能力をもっています。そのため、酢酸菌は特徴的な「Diauxie」な(2段階の)生育を示し
ます。我々人類は酢酸菌のこの「利己的」生存戦略である酸化発酵を、古くから食酢醸造とし
て、近年ではソルボース発酵やケトグルコン酸発酵などとして利用してきましたが、我々の知
らない有用な酸化発酵系が酢酸菌の酸化発酵系がまだ存在する可能性も存在しています。
この酸化発酵は、主に細胞膜表層に結合した酸化還元酵素・キノプロテインとそれにリンク
した呼吸鎖末端ユビキノール・オキシダーゼによって形成されています。このキノプロテイン
はその補欠分子族としてピロロキノリンキノンを持っていますが、その存在はabg-プロテオバ
クテリアにほぼ限定されており、ユビキノール・オキシダーゼも酢酸菌において新たに獲得さ
れたことを支持する結果が得られています。
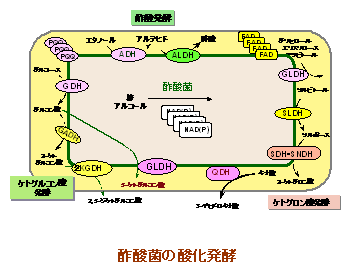
このように、酸化発酵は、この酢酸菌に特徴的な生理機能を提供する進化的に新たなシステムと考えられるようになってきました。
私たちは、このような酢酸菌の「発酵生理学」研究をすすめるとともに、酢酸菌とその酸化発酵を利用した応用研究もすすめています。それらは、酢酸発酵の耐熱化(別課題)、5-ケトグルコン酸発酵の高効率化、デヒドロシキミ酸発酵を利用したシキミ酸生産系の開発、酢酸発酵に関連して生成される菌膜多糖の有効利用、キノプロテインのバイオセンサとしての利用などです。
⇒最新の取り組み
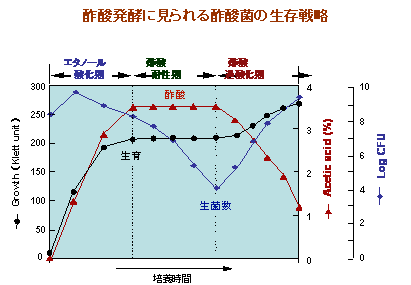
酢酸発酵の分子機構
上述した酢酸菌の「酸化発酵」の中でも、最も特徴的な反応が「酢酸発酵」です。
この酢酸発酵は細胞膜にあるアルコール酸化系呼吸鎖によるエタノールから酢酸への酸化反応に基づいています。
この反応を利用した食酢醸造は、世界各国で古くから営々と営まれてきた重要な発酵産業ですが、古来から樽や瓶の中で培地表面に薄い菌膜をつくらせる静置培養法で行なわれてきました。
今日ではタンクを用いた通気攪拌培養も行われようになってきましたが、静地培養による食酢醸造はいまでも引き続き行なわれています。
しかしながら、この酢酸発酵は、特に静置培養において、非常に不安定であり、高品質のものを安定に生産することを難しくしています。
この酢酸発酵の不安定性には、様々な要因がからんでいますが、夏場のような高温下では、菌膜が形成されないために静置培養ができず、
またタンク培養でも多量の冷却水が必要となります。加えて、生産された酢酸によって菌が死滅する現象(酢酸耐性の喪失)や生産された酢酸を消費する現象(酢酸の過酸化)が頻繁に生じます。
これらの現象は、高度な発酵制御システムを持たない中小の地場企業が、地域に根ざした伝統的な食酢の製造を行うことを困難にしています。

一方で、常に高温下にある熱帯・亜熱帯地方では、高温下で安定に酢酸発酵を行うことができないため、食酢を利用する文化が育っていません。
私たちは、このような課題に対処するため、Acetobacter 属酢酸菌に特有で酢酸発酵に重要な役割を果たす (1) エタノール酸化機構(酢酸生成能)、
(2)菌膜多糖生成機構(ペリクル形成能)、(3) 生成する高濃度酢酸に耐えて生きている本菌の酢酸耐性機構、さらに(4)酢酸発酵に有害な酢酸過酸化機構の解明を行うとともに、それらの要因と培養温度との関係についても、現在研究をすすめています。
⇒最新の取り組み
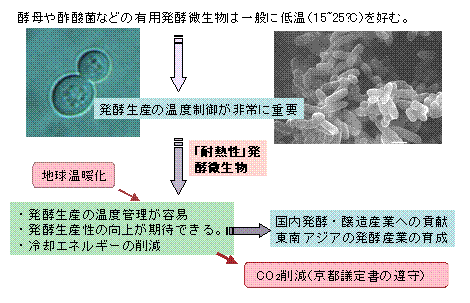
耐熱性発酵微生物(耐熱性酢酸菌)プロジェクト
酵母や酢酸菌のみならず、一般に人類に利用されている発酵微生物は比較的低温(15〜25℃)を好み、
発酵温度の上昇は菌の増殖の低下さらには発酵生産性の低下を招くことはよく知られています。
特に、昨今の地球温暖化によって、夏期には常に30度を越える高温が続くことは珍しくありません。
さらに、今日、バイオテクノロジーの東南アジアでの新興は著しく、国内微生物産業がこれらの地で、
新たに発酵産業を起こすことも多くなってきています。このような状況のもとで、比較的高温(30〜40℃)
で正常な発酵生産が行える微生物菌株の育種は非常に大きな価値をもつと考えられます。
⇒最新の取り組み
キノプロテイン脱水素酵素の構造と機能に関する研究
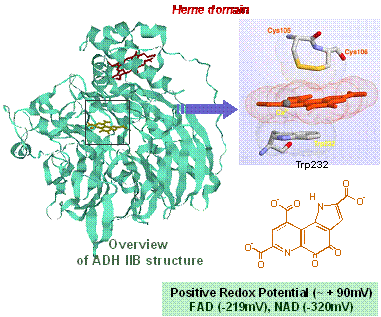
私たちは、酸化還元反応の第三番目の補酵素ピロロキノリンキノン (PQQ)をメタノール脱水素酵素(MDH)
に続いて世界で初めて細胞膜結合型アルコール脱水素酵素 (type III ADH) およびグルコース脱水素酵素 (mGDH)
に発見し、キノプロテインの反応機構の解明と生化学的基盤解析を行なってきました。
1980年代初頭からtype III ADHやmGDHに加え、可溶性のキノプロテイン、水溶性グルコース脱水素酵素(sGDH)、
PQQ に加えてへム c を補欠分子族として含む可溶性キノヘモプロテイン・アルコール脱水素酵素(type II ADH)、
酢酸菌のMDHなどの研究も開始しています。1980年代後半から1990年代前半にかけては、膜結合型キノプロテイン
とその他の呼吸鎖構成成分を精製し、それらすべてを人工膜小胞中へ再構成する手法で、キノプロテインの関与
する呼吸鎖の構造と機能を完全に明らかにしました。
⇒最新の取り組み
キノヘモプロテイン ・アルコール脱水素酵素の構造と補欠分子族